© 2002 Luluk Prihastuti
Ekowahyuni Posted:
30 November 2002
Makalah Falsafah Sains
(PPs 702)
Program Pasca Sarjana / S3
Institut Pertanian Bogor
November 2002
Dosen:
Prof Dr Ir Rudy C
Tarumingkeng (Penanggung Jawab)
Prof Dr Ir Zahrial Coto
Dr Bambang Purwantara
FENOMENA VIVIPARY LABU SIAM (Sechium edule Jacq Swartz) VARIETAS
LOKAL
DESA BARUKUPA BAWAH CIPANAS
OLEH :
LULUK PRIHASTUTI EKOWAHYUNI
A361020101/AGR
E-mail: ipb_luluk@yahoo.com
PENDAHULUAN
Latar
Belakang
Labu Siam (Sechium edule, Jacq Swartz) merupakan
tanaman sayuran dataran tinggi yang telah lama dikenal petani di Indonesia selain
bawang putih, kubis, sawi wortel, lobak dan tomat (Lingga 2001).
Labu Siam telah dikenal sebagai sayuran buah dan sekarang dikenal
sebagai sayuran pucuk (Rubatzky dan
Yamaguchi 1999). Kandungan kalori yang terdapat pada 100 g
bahan segar labu Siam buah, pucuk dan
umbi yaitu 26, 60 dan 79 kalori.
Kandungan vitamin A pada buah dan pucuk labu Siam pada 100 g bahan segar
yaitu 43 dan 4560 IU.
Berdasarkan
ciri fisiknya diduga benih labu Siam tergolong
sebagai benih rekalsitran
dengan karakteristik kadar airnya tinggi sehingga mudah terkontaminasi mikroba
dan lebih cepat mengalami kemunduran (Farrant et al. 1988). Umumnya benih
rekalsitran tidak mempunyai masa dormansi proses metabolisme perkecambahan berjalan terus (Copeland dan
McDonald 1995) bahkan benih labu Siam
dapat berkecambah ketika masih di pohon (perkecambahan dini) atau bersifat vivipary.
Sifat tanaman yang mirip dengan labu Siam diantaranya adalah tanaman
species mangrove (Tomlinson 1998). Labu Siam tidak tahan disimpan
sebagai benih lebih dari satu bulan
sejak berkecambah di pohon karena tidak memiliki masa dormansi sehingga diduga labu Siam termasuk
dalam rekalsitran tinggi (highly
rekalsitran). Hal ini menunjang
pendapat Farrant et al. (1988) mengenai beberapa karakteristik
benih rekalsitran.
Perbanyakan
tanaman labu Siam selama ini dilakukan secara generatif dengan penanaman buah
yang matang di batang dan telah berkecambah (Rubatzky dan Yamaguchi 1999). Buah yang dipakai sebagai benih merupakan
panenan pertama, terletak pada batang utama, mempunyai ciri-ciri fisik yang baik, dan kotiledon dalam keadaaan
sehat.
Perbanyakan
tanaman dengan cara vegetatif adalah dengan stek yang telah berakar sempurna
yang diperoleh dari batang yang muda
namun cara ini jarang dilakukan karena produksi dan produktivitas
buahnya rendah. Rukmana (1999)
menambahkan bahwa benih yang baik
dihasilkan dari pohon induk yang baik. yakni tanaman tumbuh subur normal, berbuah lebat stabil, umur tanaman
cukup dan keadaan tanaman sehat tidak berpenyakit atau terserang hama. Benih yang akan dijadikan bibit harus
dipilih benih yang baik, bermutu, buah berumur tua, dan bentuknya normal,
terletak di bagian tengah batang atau pada batang pokok, ukuran benih seragam,
benih tidak diserang hama dan penyakit.
Selama ini
benih labu Siam dikembang biakkan dalam bentuk buah yang sudah berkecambah dan
sehat pada umur 42 hari setelah anthesis (HSA), buah telah berakar dan
berkecambah sepanjang 2-4 cm dengan daun sepasang. Benih labu Siam yang digunakan untuk perbanyakan tanaman beratnya
rata-rata 300-400 gram dengan kondisi voluminous dan resiko kerusakan yang
tinggi. Transportasi benih dari daerah
pertanaman labu Siam yang menyebar ke seluruh wilayah Indonesia merupakan hal
yang sulit. Menurut Lingga (2001) kebutuhan benih per hektar
untuk labu Siam adalah sekitar 650 benih/ha. Tahun 2001 luas areal pertanaman
baru untuk labu Siam 29.223 ha maka total kebutuhan benih sekitar 18.994.950
benih.
Penelitian
mengenai kandungan gizi kegunaan dan jumlah species labu Siam telah banyak
dilakukan di Luar Negeri seperti di Negara Amerika Tengah. Masih banyak permasalahan yang belum
diketahui pada benih labu Siam
khususnya mengenai fenomena vivipary labu Siam varietas lokal daerah Cipanas
yang merupakan daerah sentra labu Siam.
Penelitian
ini mengamati fenomena vivipary labu
Siam melalui analisis kandungan hormon ABA
(asam absisat) dan hormon IAA (asam indole-3 asetic),
Diharapkan
hasil penelitian ini memberikan informasi tambahan tentang fenomena vivipary benih labu Siam.
Tujuan
Penelitian
Penelitian ini dilaksanakan dengan tujuan sebagai berikut :
Memperoleh informasi tentang sifat vivipary labu Siam berdasarkan analisis kandungan ABA dan IAA
Hipotesis
Penelitian
Terdapat
penurunan kandungan hormon ABA dan
peningkatan kandungan hormon IAA pada
fenomena vivipary benih labu Siam.
 TINJAUAN
PUSTAKA
TINJAUAN
PUSTAKA
Labu Siam (Sechium edule, Jacq
Swartz)
Labu Siam (chayote) merupakan salah satu tanaman sayuran
dataran tinggi di Indonesia. Buah,
pucuk, akar dan umbi labu Siam semua bisa dikonsumsi. Menurut Engels (1983) di
Papua Nugini pucuk umbi dan buah digunakan sebagai makanan semua jenis ternak. Tanaman labu Siam mempunyai prospek sebagai dietary
food, karena mempunyai kandungan
kalori yang rendah dan digunakan sebagai makanan penambah rasa. Bijinya berbentuk seperti kacang yang
mengandung sumber protein. Pucuk
khususnya kaya akan vitamin A, B dan
C. Di Indonesia tidak ada statistik secara tersendiri data labu Siam selalu dikombinasi dengan
semua tanaman labu (Biro Pusat Statistik
1998).
Dalam
produksi dan perdagangan Internasional, labu Siam adalah termasuk 5 (lima)
jenis sayuran komersial yang penting di Brazil. Ini merupakan Informasi penting
bagi Indonesia karena di Indonesia labu
Siam sangat cocok tumbuh dan berproduksi terus sepanjang tahun. Menurut Rukmana (1999) tanaman labu Siam
dalam pertumbuhan dan perkembangannya adalah tanaman hijau sepanjang
tahun. Tanaman ini direkomendasikan untuk diperbaiki paling sedikit tiga tahun sekali, terutama
apabila terserang penyakit dan untuk menghindari serangan penyakit.
Syarat
tumbuh bagi tanaman labu Siam adalah
kelembaban relatif tinggi (80-85%) curah hujan tahunan paling sedikit
1500 – 2000 mm terdapat Irigasi dan temperatur rata-rata adalah 20 – 250C
(dengan batas 12 - 28 0C).
Pertumbuhan terbaik bagi labu Siam adalah pada ketinggian 300 m – 2000 m
di atas permukaan laut (dpl) dengan tanah yang berdrainase baik. Labu Siam apabila ditanam di dataran rendah
maka tidak bisa berproduksi menghasilkan buah (Engels 1983).
Pembungaan
dimulai 1 – 2 bulan sesudah perkecambahan dan pembungaannya menurut Rukmana (1999) berlimpah sepanjang
tahun. Bunga tanaman labu Siam adalah
menyerbuk silang tetapi self compatible dan
berumah satu yakni bunga jantan dan betina terdapat dalam satu tanaman. Bunga jantan mirip dengan bunga betina
tetapi berukuran kecil dan tiap tandan terdiri banyak kuntum terletak dalam
satu batang.
Buah
terbentuk tiga bulan setelah ditanam.
Buah yang diproduksi jumlahnya ratusan per pohon per tahun. Perkecambahan bisa terjadi ketika buah
berada di pohon. Fenomena ini disebut vivipary mirip seperti species mangrove.
Labu Siam varitas lokal Cipanas tidak bisa disimpan sebagai benih
lebih dari satu bulan sejak berkecambah di pohon, karena benih tidak memiliki
masa dormansi. Selama ini penyimpanan
labu Siam adalah dalam bentuk buah.
Engels (1983) mengemukakan pula bahwa penyimpanan atau pemeliharaan
plasma nutfah labu Siam harus dalam bentuk tanaman hidup atau kultur jaringan
di bawah kondisi kelembaban rendah.
Koleksi plasma nutfah labu Siam di seluruh dunia dihasilkan oleh Chapingo Regional Centre (Mexico) dan
beberapa perusahaan lain. Buah labu
Siam berbentuk bulat sampai agak lonjong menyerupai buah alpukat dan mengandung
tangkai buah. Struktur buah
terdiri-dari kulit buah yang tipis dan berduri jarang, daging buah yang amat
tebal berbiji tunggal, daging buah banyak mengandung air dan getah. Getah labu
Siam berkhasiat sebagai obat penurun panas badan.
Bijinya
berbentuk panjang atau lonjong pipih berkeping dua. Akan ditelaah apakah biji tanpa
buahnya (benih) dapat digunakan untuk perbanyakan tanaman secara
generatif.
Fenomena vivipary
Vivipary adalah perkecambahan dini yang terjadi karena embrio
yang dihasilkan berasal dari reproduksi sexual normal tidak mempunyai masa
dormansi, pertumbuhan pertama kecambah keluar melalui kulit benih dan
selanjutnya keluar melalui buah ketika tanaman masih berada di batang tanaman
induknya. Proses ini terjadi pada
beberapa species tanaman diantaranya
labu Siam mangrove beberapa kultivar buah seperti citrus dan ophiorhiza. Tanaman vivipary banyak ditemukan di daerah wetlands (basah).
Hal yang
menarik bahwa fenomena vivipary, bisa
diamati secara morfologi, ekologi maupun fisiologi. Fisiologi dari vivipary adalah
bervariasi karena adanya kondisi konsentrasi garam di dalam tanah (media),
aktivitas respirasi dan distribusi enzym maupun hormon.
Penelitian
ini akan mengamati fenomena vivipary
berdasarkan distribusi hormon di dalam perkembangan tanaman labu Siam. Penelitian ini berarti mengamati fenomena vivipary dari aspek fisiologinya. Menurut Salisbury dan Ross (1995) yang
dimaksud hormon tumbuhan adalah senyawa organik yang di sintesis di salah satu
bagian tumbuhan dan dipindahkan ke bagian lain dan pada konsentrasi yang sangat
rendah mampu menimbulkan suatu respon fisiologi. Respon pada organ sasaran tidak selalu bersifat memacu, karena
suatu proses pertumbuhan dan diferensiasi kadang malah menghambat misalnya ABA (Inhibitor). Hormon khas pada tumbuhan karena effektif
berkerja pada konsentrasi yang amat rendah.
Hormon sering effektif pada konsentrasi 1 mikromolar sehingga senyawa
kimia lain yang aktif pada konsentrasi tinggi bukan hormon misalnya vitamin dan
sukrosa.
Salisbury
dan Ross (1995) menambahkan hormon yang pertama kali ditemukan adalah auksin.
Auksin endogen yaitu IAA (Indol
Acetic Acid) ditemukan pada tahun
1930-an bahkan saat itu hormon mula-mula dimurnikan dari air seni. Karena semakin banyak hormon ditemukan maka
efek serta konsentrasi endogennya dikaji.
Hormon pada tanaman jelas mempunyai ciri : setiap hormon mempengaruhi
respon pada bagian tumbuhan, respon itu bergantung pada species, bagian
tumbuhan, fase perkembangan, konsentrasi hormon, interaksi antar hormon, yang
diketahui dan berbagai faktor lingkungan yaitu cahaya, suhu, kelembaban, dan
lainnya.
Hormon ABA (Asam absisat)
Semua
jaringan tanaman terdapat hormon ABA
yang dapat dipisahkan secara kromatografi Rf
0.9. Senyawa tersebut merupakan
inhibitor B –kompleks. Senyawa ini mempengaruhi proses pertumbuhan,
dormansi dan absisi. Beberapa peneliti
akhirnya menemukan senyawa yang sama yaitu asam
absisat (ABA). Peneliti tersebut yaitu Addicott et al dari California USA pada tahun
1967 pada tanaman kapas dan Rothwell serta Wain pada tahun 1964 pada tanaman
lupin (Wattimena 1992).
Menurut Salisbury dan Ross (1995) zat
pengatur tumbuhan yang diproduksi di dalam tanaman disebut juga hormon tanaman.
Hormon tanaman yang dianggap sebagai hormon stress diproduksi dalam jumlah
besar ketika tanaman mengalami berbagai keadaan rawan diantaranya yaitu ABA.
Keadaan rawan tersebut antara lain kurang air, tanah bergaram, dan suhu dingin atau panas. ABA membantu tanaman
mengatasi dari keadaan rawan tersebut.
ABA adalah seskuiterpenoid berkarbon 15,
yang disintesis sebagian di kloroplas dan plastid melalui lintasan asam
mevalonat (Salisbury dan Ross 1995).
Reaksi awal sintesis ABA sama
dengan reaksi sintesis isoprenoid
seperti gibberelin sterol dan karotenoid. Menurut Crellman (1989) biosintesis ABA pada sebagian besar tumbuhan terjadi secara tak langsung melalui peruraian karotenoid tertentu (40 karbon) yang ada
di plastid. ABA pergerakannya dalam tumbuhan sama dengan pergerakan gibberelin yaitu dapat diangkut secara
mudah melalui xilem floem dan juga sel-sel parenkim di luar berkas
pembuluh.
Hormon IAA (asam indol- 3 asetat)
Istilah auksin pertama kali digunakan oleh Frist
Went seorang mahasiswa PascaSarjana di negeri Belanda pada tahun 1926 yang kini
diketahui sebagai asam indol-3 asetat atau
IAA (Salisbury dan Ross 1995).
Senyawa ini terdapat cukup banyak di ujung koleoptil tanaman oat ke arah cahaya. Dua mekanisme sintesis IAA yaitu pelepasan gugus amino dan gugus karboksil akhir dari
rantai triphtofan. Enzim yang paling aktif diperlukan untuk
mengubah tripthofan menjadi IAA terdapat di jaringan muda seperti
meristem tajuk, daun serta buah yang sedang tumbuh. Semua jaringan ini kandungan IAA
paling tinggi karena disintesis di
daerah tersebut.
IAA terdapat di akar pada konsentrasi
yang hampir sama dengan di bagian tumbuhan lainnya (Salisbury dan Ross 1995).
IAA dapat memacu pemanjangan
akar pada konsentrasi yang sangat rendah.
IAA adalah auksin endogen atau
auksin yang terdapat dalam
tanaman. IAA berperan dalam aspek pertumbuhan dan perkembangan tanaman yaitu
pembesaran sel yaitu koleoptil atau batang penghambatan mata tunas samping,
pada konsentrasi tinggi menghambat pertumbuhan mata tunas untuk menjadi tunas
absisi (pengguguran) daun aktivitas dari kambium dirangsang oleh IAA pertumbuhan akar pada konsentrasi tinggi dapat menghambat
perbesaran sel-sel akar.
Penelitian
IAA oleh Gregorio et al (1995) pada
embrio, endosperma, dan integumen benih Sechium
edule (labu Siam) pada umur 23, 27, 33, dan 37 hari setelah anthesis adalah
sebagai berikut: 1) jumlah IAA pada embrio pada umur tersebut berturut-turut
1.67%, 2.08%, 3.40 % dan 3.29 %, 2) Jumlah IAA pada endosperma berturut-turut
20.45%, 25.72%, 30,40%, dan 52.22% dari total IAA, dan 3) Jumlah IAA pada
integumen adalah 8.44%, 9.32%, 8.76% dan 8.04%, dan 4) Jumlah IAA total ( IAA terikat maupun IAA bebas) cenderung
meningkat sejalan dengan meningkatnya kemasakan benih labu Siam.
METODOLOGI
Penelitian dilakukan di empat lokasi yaitu : 1) kebun sayur Pacet
desa Barukupa Bawah kabupaten Cianjur Jawa Barat dan 2) laboratorium Balai
Penelitian Bioteknologi Cimanggu Bogor .
Penelitian
dimulai bulan Juli 2000 sampai dengan bulan Juni 2001. Penelitian tentang fenomena vivipary labu Siam dengan cara manganalisa
kandungan hormon ABA dan IAA
Fenomena vivipary dengan menganalisis kandungan
hormon ABA dan IAA
Pengamatan fenomena vivipary benih labu Siam dilakukan
dengan menggunakan empat (4) tingkat kemasakan dengan 3 ulangan (3 buah) yaitu
:
M0 (14 HSA),
M1 (21 HSA), M2 (28 HSA) yaitu berturut-turut stadia 9,10,11
(diduga merupakan saat masak fisiologis benih) dan 13 yaitu M3 (42
HSA).
1. Analisis kandungan ABA
Analisis ABA menggunakan metode Robertson dan Synder et al. (1987) dengan alat High Performance Liquid
Chromatography. Tahap analisis mencakup penyimpanan ekstrak labu Siam dengan tiga ulangan (3 buah labu
Siam). Ekstrak dari jaringan
benih labu Siam yaitu bagian embrio dan kotiledonnya disimpan
dalam nitrogen cair. Kemudian di purifikasi dengan larutan
methanol : akuades : asam asetat ( 50 : 49 : 1, v/v). Penetapan kandungan ABA, larutan contoh disuntikkan ke alat High Performance Liquid
Chromatographi. Fase diam yang
digunakan adalah kolom
C 18 sedangkan fase cair
adalah metanol : asam asetat : akuades. Detektor dengan l 260 nm sedang kecepatan
alir fase gerak adalah 1 ml/ menit suhu detektor 25 0 C dengan attenuasi 0.02.
2.
Analisis kandungan IAA
Analisis
kandungan IAA ini menggunakan metode
Sandberg et al. (1987).
Tahap analisis mencakup
penyimpanan ekstrak labu Siam dengan 3 ulangan (3 buah labu Siam). Ekstrak dari jaringan benih labu Siam yaitu
bagian embrio dan kotiledonnya disimpan dalam larutan metanol 0.3 g/ml yang mengandung
0.02 % natrium dietilkarbamat, selama 2 jam. Ekstrak metalonat dipurifikasi
dengan kromatografi XAD, kemudian dicuci 5 ml etil asetat /hexane ( 3:1, v/v) , dan disuntikkan pada alat High Performance Liquid Chromatographi.
HASIL DAN PEMBAHASAN
Fenomena vivipary
dengan menganalisis kandungan hormon ABA
dan IAA
Bagan berikut menunjukkan kerja
beberapa hormon yaitu hormon
yang memacu pertumbuhan dan menghambat pertumbuhan (inhibitor) dalam mempengaruhi perkecambahan dan
dormansi biji.
Fenomena vivipary selain pada labu Siam banyak juga
terdapat pada tanaman mangrove famili
Rhizophoraceae dan beberapa spesies
buah-buahan. Tan dan Rao (1981)
menyatakan vivipary banyak terdapat
pada tanaman di lahan basah (wetlands). Ditambahkan oleh Tan dan Rao (1981) bahwa
terdapat pula vivipary palsu pada
famili Rubiaceae inflourecense pada
tanaman Agave dan Poa alpina tanpa didahului oleh proses
seksual karena hal ini tidak terjadi pada tanaman yang berbunga. Benih labu Siam perkecambahan terus terjadi
ketika di pohon induknya.
Kondisi
fenomena vivipary pada mangrove menyebabkan tanaman ini
mempunyai daya adaptasi yang tinggi terhadap lingkungannya (kadar garam tinggi,
sinar matahari menyengat, angin yang keras).
![]()
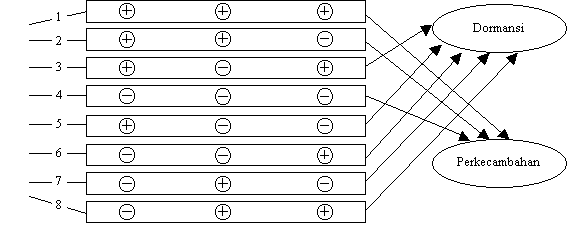
Gambar 1. Model fungsi seleksi dari hormon terhadap perkecambahan dan
dormansi
KESIMPULAN DAN SARAN
KESIMPULAN
Fenomena vivipary terjadi karena kandungan ABA semakin menurun dan kandungan IAA semakin meningkat dengan semakin
meningkatnya stadia kemasakan benih.
Fenomena vivipary tidak hanya
dipengaruhi oleh kandungan ABA saja
tetapi diduga lebih banyak merupakan hasil kerjasama dengan fitohormon lainnya
seperti auksin endogen (IAA), giberrelin, sitokinin dan etilen.
SARAN
Perlu
dilakukan penelitian lebih lanjut untuk mengetahui dengan lebih baik tentang
fenomena vivipary benih labu Siam
pada peneliti selanjutnya dianjurkan untuk menganalisis kandungan fitohormon
lainnya giberellin sitokinin, serta
ethilen.
DAFTAR
PUSTAKA
Adimargono. 1997.
Recalcitant seeds, Identification and storage. Thesis. Larenstein International Agricultura Collage. Deventer.
Anonim.
1995. Produksi tanaman sayuran dan buah-buahan
semusim di Jawa. Biro Pusat
Statistik. Jakarta Indonesia. 3 hal.
Berjak
P and N W Pammenter. 1994. Recalcitrance is not an all or nothing
situation. Seed Sci. Res 4 : 263-264.
Bewley
J .D and M Black. 1994. Seeds:
physiology of development and germination. 3 rd Edition Plenum Press. New York and London. 445p.
Bonner F T.
1996. Response to drying of
recalcitrant seed of Quercus nigra L. Ann.
Bot 78 : 181 – 187.
Ceccarelli
N and Lorenzi R. 1992. Role of cotyledon and gibberellines in the
early growth of Sechium edule. Sw plantlets. Journal of palnt physiology.
Italy. 5 hal.
Chandler
P M. 1988. Hormonal regulation of gene expression in the “Slender” mutant of
barley (Hordeum vulgare L). Planta.
175 : 115 –120.
Chin H
F and E H Roberts. 1980. Recalcitrant crop seeds. Kuala Lumpur, Malaysia.
Tropical Press SDN. BHD.
------------,
B. Chrishnapillay and P.C. Standwood
1989. Recalsitrant Vs Orthodox
Seeds, P. 15-22. In Seed Moisture. CSSA Special Publication Number 14. Crop Science Society of America Madison,
Winsconsin, USA.
Copeland
LO dan Mc Donald M B. 1976. Principles
of seed science and technology.
Macmilan Publs Cry New York and Callier Macmillan Publ London . 321 p.
Creelman
R A, Barendse G W M, Galston A W, Robertson J M, and Thompson G A. 1987.
High performance liquid chromatography in Plant Sciences. Eddited by H.F Linkens dengan j F
Jackson. Aspringer Verlag Berlin
Heidelberg New York Paris Tokyo . hal 79-88.
Duke N
C, Bunt J S, and Williams W T.
1984. Observations of the floral
and vegetative phenologies of North estern Australian mangroves. Australia. Journal Botany. 32 : 87-89.
Engels
J M M. 1983 . Variation in Sechium edule.
SW. In Central America. Journal of
the American Society for Horticultural Science. 108 hal : 706 – 710.
Engels
J M M. 1984. Chayote : a little known. Central American Crop. FAO/IBPGR Plant
Genetic Resources Newsletter . New
York.
Gill A
M and Tompson P B. 1977. Studies of growth of red mangrove
(Rhizophora mangle L).4. The adult root system. Biotropica 9(3) : 145- 155.
Gregorio
S, Passerini P, Picciarelli P, and Ceccarelli N. 1995. Free and conyugated indole-3- acetic acid in Developing seeds of Sechium
edule, Jacq Swartz. Journal of
Plant Physiologi Italy ; 145
; 5-6, 736 – 790.
Hopkins W G.
1995. Introduction to plant
physiology. New York, Brisbane, Toronto, Singapore. John Willey and Sons, Inc.
Johri B M.
1984. Embryology of angiosperms.
New York Tokyo. Springer Verlag Berlin
Heidelberg .
Lingga.
P. 2001. Panduan seminar dan peluncuran buku retrospeksi perjalanan
Industri benih di Indonesia. Bogor.
P.T. Sang Hyang Seri & Laboratorium Ilmu & Teknologi Benih Jurusan Budidaya Pertanian Faperta
Institut Pertanian Bogor.
Rubatzky
dan Yamaguchi. 1999. Sayuran Dunia 3:
Prinsip, Produksi, dan Gizi. Bandung.
Penerbit Institut Teknologi Bandung .
Rukmana R. 1998. Budidaya Tanaman Labu Siam (Sechium edule,. Jacq Swartz).
Jogjakarta. Penerbit Kanisius Jogjakarta.
Salisbury
F B, dan Ross C W. 1995. Fisiologi Tumbuhan. Bandung.
Penerbit ITB Bandung.
Sandberg
G, Crozier A, Chen Ch M, Hartley R D, and Smith L A. 1987. High Performance Liquid Chromatography in Plant
Sciences. Eddited by H.F Linkens dengan
J F Jackson. Aspringer Verlag Berlin
Heidelberg New York Paris Tokyo .
Synder
and Robertson J M. 1987. The determination of absisic acid by High performance
liquid chromatography, In . H.H. Linshens and J.F. Jackson (Eds). High
performance liquid chromatography in Plant sciences. (Eds). High Performance
liquid chromatography in plant sciences. Springer- Verlag.Berlin. Heidelberg.
New York. London. Paris. Tokyo. p:
52 – 71
Tomlinson
P B. 1998. The Botany of Mangroves. Cambridge. London, New York,
Melborne, Sydney. Cambridge University
Press.
Wang
Y, S S Li, J X He, and J R Fu. 1977.
Protective enzymes against activated oxygen in recalcitrant wampee
(Clausena lansium) seeds during dessication.
p. 147-152. In. A G Taylor and Xue Ling Huang (Eds). Progress in seed research.
Conference proceedings of the 2 nd ICSST. China.
Wattimena G A. 1998. Zat Pengatur Tumbuh Tanaman.
Bogor. Pusat Antar Universitas. Institut
Pertanian Pertanian Bogor.